
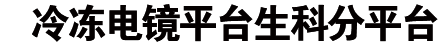

前沿速递 | 北京大学郭强团队首次解析过氧化物酶体中尿酸氧化酶的超分子组装机制,为代谢酶稳态调控提供新见解
转自LTS预印本平台
在哺乳动物的肝细胞中,过氧化物酶体是尿酸氧化酶的主要分布场所,在嘌呤代谢中起着关键作用[1]。长期以来,电镜观察发现其内部存在致密的“类晶体核心”结构,被推测主要由尿酸氧化酶(UOX)组成[2]。传统体外研究往往将酶置于稀释缓冲液中,无法还原细胞内的拥挤环境,因此难以捕捉这种天然的高阶组装状态[3]。因此这一结构的精确分子架构、组装机制和生理功能始终是未解之谜。
北京时间2026年4月18日,北京大学郭强、陈晓伟团队在浪淘沙预印本平台发布了标题为“A conserved helix-tiling architecture of urate oxidase organizes metabolism in peroxisomes”的最新研究进展(图1)。依托原位冷冻电镜断层扫描技术,首次在小鼠肝细胞中解析了尿酸氧化酶(UOX)独特的“螺旋平铺”高阶组装结构,揭示了哺乳动物细胞中代谢酶应对严酷的细胞内环境的独特生存策略。该结构由UOX同源四聚体通过保守界面自组装形成多孔片层支架,在不改变酶催化活性的前提下,大幅提升了酶对热、蛋白酶解和氧化应激的抗性。

图1. 成果发布在浪淘沙预印本平台(Langtaosha.org)
破解“类晶体”之谜:独特的螺旋平铺架构
研究团队首先对小鼠肝脏组织进行原位冷冻电镜成像,在过氧化物酶体中捕捉到了具有高度周期性的超分子组装结构。通过亚断层平均技术和单颗粒分析技术,团队将该结构的分辨率推进至3.64埃,成功鉴定其组成成分为尿酸氧化酶(UOX)。
解析结果显示,UOX的基本功能单元是同源四聚体,这些四聚体通过三个保守的分子界面相互作用,自组装成右螺旋的空心纤维。每个纤维的内径约3.5纳米,外径约21纳米,相邻纤维以约144°的夹角侧向平铺延伸,最终形成单分子层的多孔片层支架。这一结构完美解释了为什么传统观察会将其误认为“晶体”——其内部具有严格的周期性排列,但纤维的五重对称性违背了晶体的空间填充规则,因此不会形成致密的块状晶体,反而保留了大量孔隙通道。
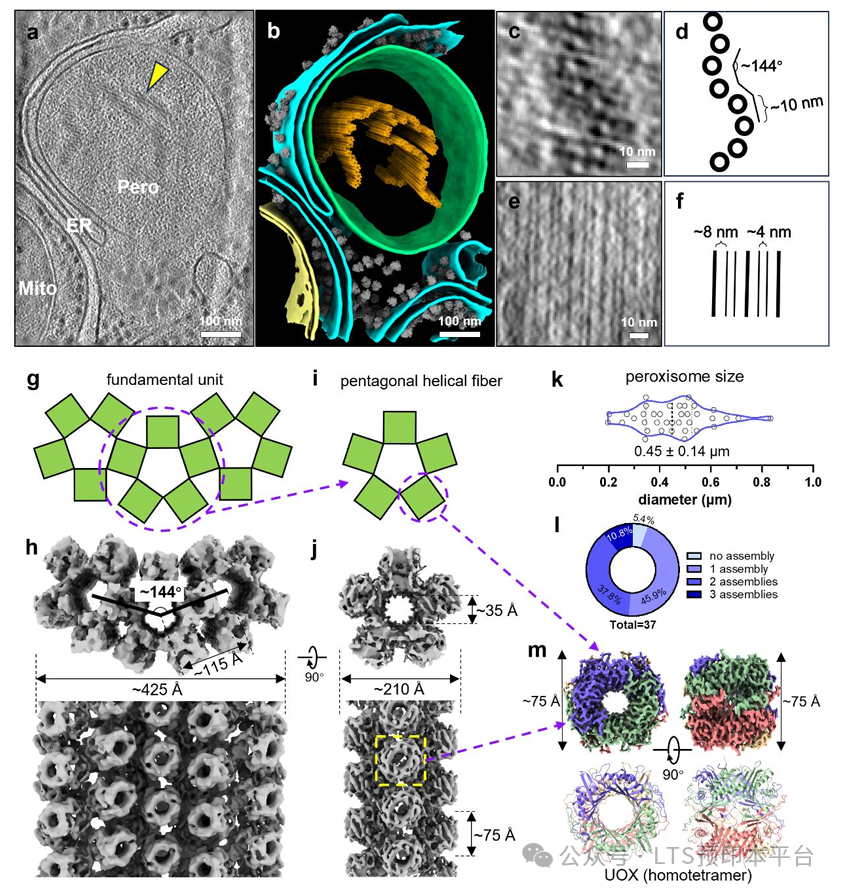
图2:小鼠肝脏过氧化物酶体内的超分子尿酸氧化酶组装体
这一“螺旋平铺”架构极具巧思:一方面,紧密的分子间相互作用赋予结构极高的机械稳定性;另一方面,多孔性使得底物尿酸和产物可以自由扩散,而所有四聚体的催化位点都通过相邻纤维间的界面B朝向孔隙,确保酶的活性中心完全暴露,不会因组装而被遮蔽。
揭示组装机制:保守界面介导的层级自组装
为了阐明组装的分子机制,团队对UOX四聚体间的相互作用界面进行了精细解析。研究发现,三个界面分工明确:界面C是介导螺旋纤维纵向延伸的核心,由Arg202与Asn148之间的静电相互作用以及亮氨酸、异亮氨酸组成的疏水补丁共同稳定;界面A负责纤维间的侧向交联;而界面B则紧邻催化位点,没有明显的相互作用,保证底物可自由结合。
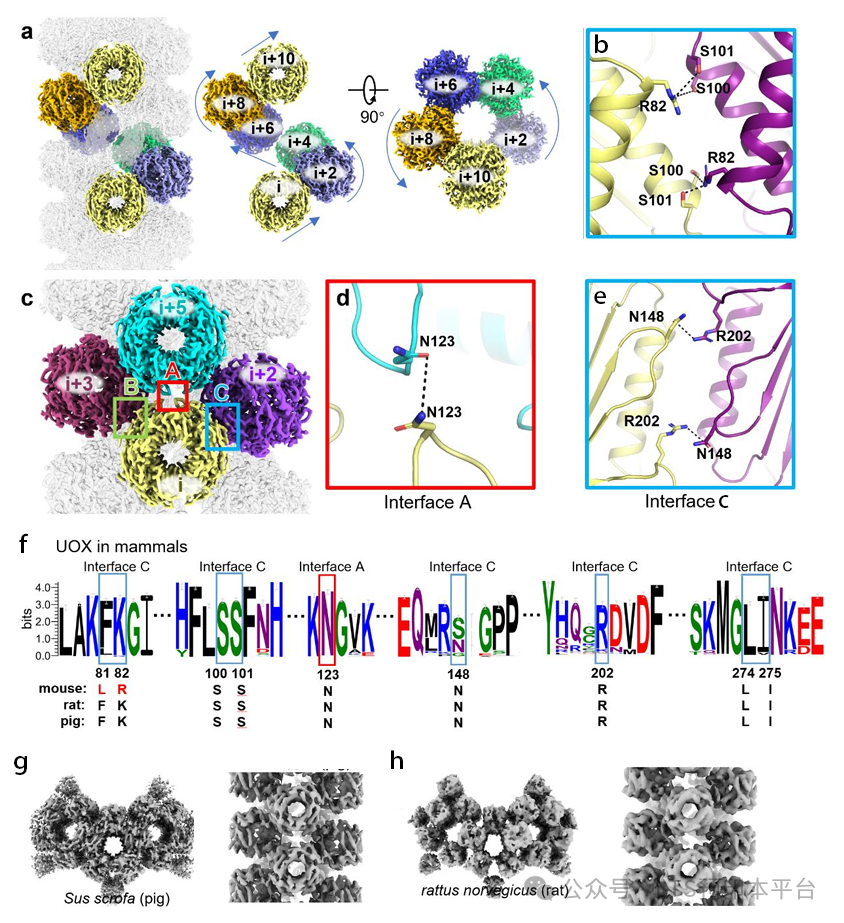
图3. UOX组装体机制
通过对不同哺乳动物的UOX序列进行比对,团队发现介导组装的关键残基在所有表达UOX的哺乳动物(小鼠、大鼠、猪、牛等)中高度保守。对猪和大鼠肝脏的原位结构验证也证实,这一螺旋平铺结构是哺乳动物的普遍特征。而在鱼类、两栖类等更低等的脊椎动物中,这些关键残基发生了变异,其UOX以可溶性形式存在于过氧化物酶体中,提示这一组装是哺乳动物特有的进化适应。
新的功能调控:提升稳定性而不影响活性
以往发现的代谢酶高阶组装往往通过变构效应调控催化活性,而UOX的组装却呈现出完全不同的逻辑。体外酶活实验显示,组装状态的UOX与游离四聚体的催化活性几乎没有差异,分子动力学模拟以及结构信息也证实催化过程和组装不会引起蛋白构象变化。在小鼠体内实验中,研究人员通过定点突变破坏UOX的组装界面,突变体仍能保持正常催化活性,小鼠的基础血清尿酸水平也与野生型无异。
那么这种组装的生理意义是什么?答案在于过氧化物酶体的特殊环境——UOX催化反应会产生大量过氧化氢,这一强氧化剂极易损伤蛋白。实验证实,组装状态的UOX相比游离四聚体,解链温度提升了约5℃,对蛋白酶K的抗性有所提高,而在氧化应激条件下的氧化修饰水平则有所降低。这意味着,通过组装成螺旋平铺结构,UOX在不牺牲催化效率的前提下,大幅提升了在高活性氧环境中的稳定性,从而实现长期、持续的尿酸代谢功能。
图4. UOX组装体形成的功能影响
研究团队将这一策略总结为“分子鲁棒性优化”:在恶劣的细胞微环境中,酶不需要通过变构调节活性,而是通过自组装提升自身抗逆性,保证代谢流的稳定供给。这一发现突破了传统关于代谢酶组装的功能认知,丰富了细胞在拥挤环境中优化生化反应的策略库。
应用前景:为尿酸酶药物设计提供新思路
人类在进化过程中丢失了UOX基因,因此尿酸是嘌呤代谢的终产物,血液中尿酸浓度过高会引发痛风、肾结石等疾病。目前临床使用的尿酸酶药物(如拉布立酶、普瑞凯希)均来自真菌或猪-狒狒嵌合体,存在稳定性差、半衰期短、免疫原性强等缺陷。
该研究揭示的UOX组装机制为下一代尿酸酶药物的设计提供了新方向:通过改造人源化UOX的组装界面,使其在生理条件下自发形成稳定的高阶组装体,有望在保持低免疫原性的同时,显著提高酶的稳定性和体内半衰期,从而改善现有药物的不足。此外,这种“保持活性、提升稳定”的组装策略,也可扩展至其他工业酶或治疗性酶的改造,通过理性设计组装界面,获得性能更优的酶制剂。
北京大学团队的这项工作,不仅解答了关于过氧化物酶体核心结构的长年疑问,也展示了原位结构生物学在揭示生命活动机制方面的能力。生命系统的精巧往往体现在微妙的适应策略中:进化未必让单个酶分子更“强”,而是使其通过有序组装,在复杂环境中以更稳健的方式实现功能优化。这一发现为理解细胞内的代谢组织逻辑提供了新的视角,也为相关生物工程应用开辟了可能的路径。
原文链接:
https://langtaosha.org.cn/lts/en/preprint/view/189
参考文献:
1.Noguchi, T., Takada, Y., and Fujiwara, S. (1979). Degradation of uric acid to urea and glyoxylate in peroxisomes. Journal of Biological Chemistry 254, 5272-5275. 1493 10.1016/S0021-9258(18)50590-1.
2.Kahn, K., Serfozo, P., and Tipton, P.A. (1997). Identification of the True Product of the Urate Oxidase Reaction. Journal of the American Chemical Society 119, 5435-5442. 10.1021/ja970375t.
3.Carcia-Seisdedos, H., Empereur-Mot, C., Elad, N., and Levy, E.D. (2017). Proteins evolve on the edge of supramolecular self-assembly. Nature 548, 244-247. 10.1038/nature23320.